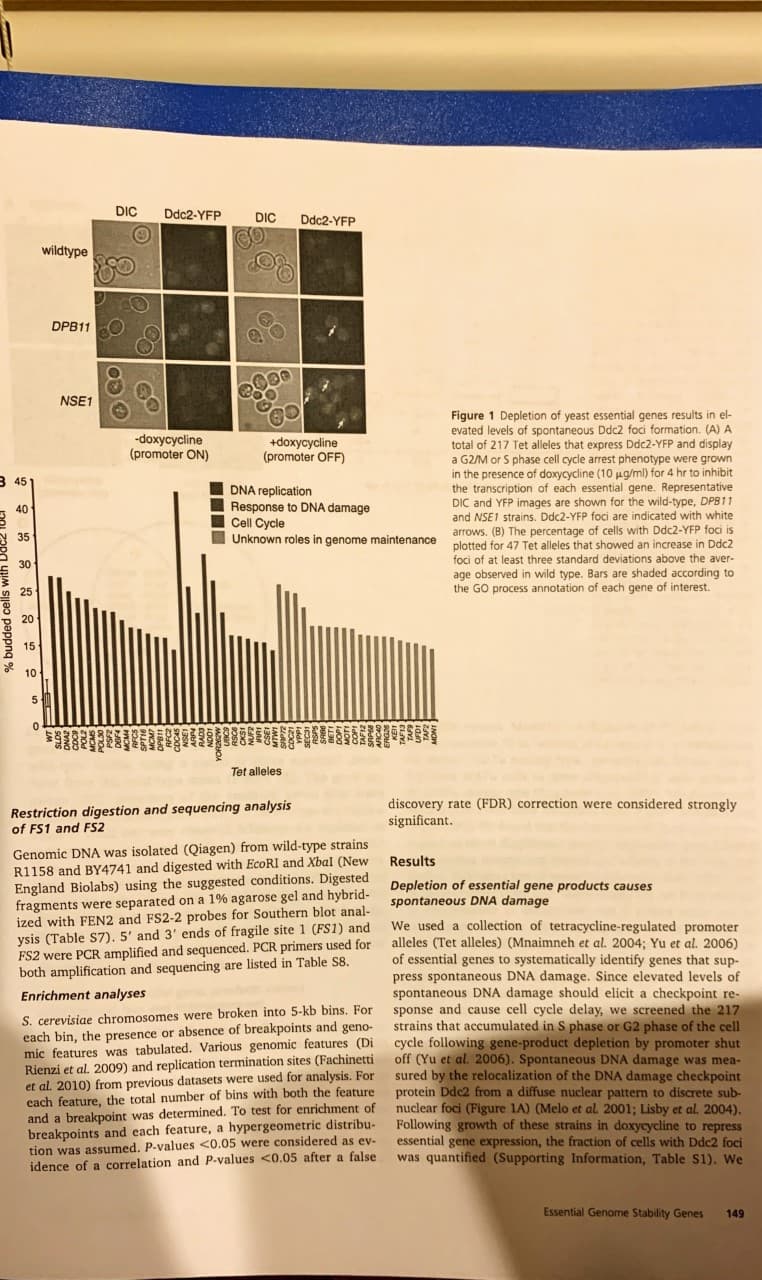
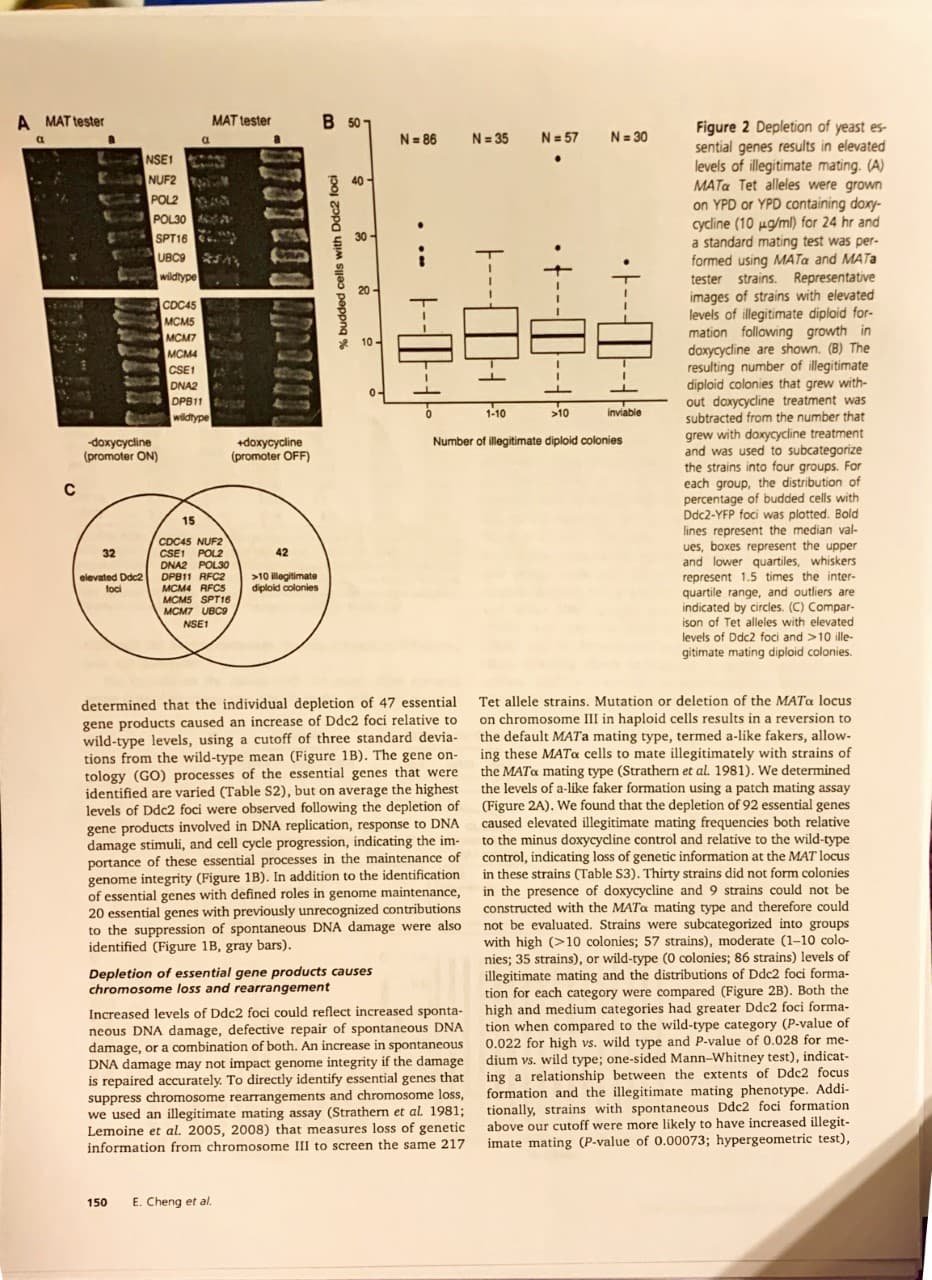
DIC Ddc2-YFP DIC Ddc2-YFP wildtype DPB11 NSE1 Figure 1 Depletion of yeast essential genes results in el- evated levels of spontaneous Ddc2 foci formation. (A) A total of 217 Tet alleles that express Ddc2-YFP and display a G2/M or S phase cell cycle arrest phenotype were grown in the presence of doxycycline (10 ug/ml) for 4 hr to inhibit the transcription of each essential gene. Representative DIC and YFP images are shown for the wild-type, DPB11 and NSE1 strains. Ddc2-YFP foci are indicated with white -doxycycline (promoter ON) +doxycycline (promoter OFF) 3 45 DNA replication Response to DNA damage Cell Cycle Unknown roles in genome maintenance arrows. (B) The percentage of cells with Ddc2-YFP foci is plotted for 47 Tet alleles that showed an increase in Ddc2 foci of at least three standard deviations above the aver- observed in wild type. Bars are shaded according to the GO process annotation of each gene of interest 20 10 5 Tet alleles discovery rate (FDR) correction were considered strongly significant. Restriction digestion and sequencing analysis of FS1 and FS2 Genomic DNA was isolated (Qiagen) from wild-type strains R1158 and BY4741 and digested with EcoRI and Xbal (New England Biolabs) using the suggested conditions. Digested fragments were separated on a 1% agarose gel and hybrid- ized with FEN2 and FS2-2 probes for Southern blot anal- ysis (Table S7). 5' and 3' ends of fragile site 1 (FS1) and FS2 were PCR amplified and sequenced. PCR primers used for both amplification and sequencing are listed in Table S8. Results Depletion of essential gene products causes spontaneous DNA damage We used a collection of tetracycline-regulated promoter alleles (Tet alleles) (Mnaimneh et al. 2004; Yu et al. 2006) of essential genes to systematically identify genes that sup- press spontaneous DNA damage. Since elevated levels of spontaneous DNA damage should elicit a checkpoint re- sponse and cause cell cycle delay, we screened the 217 strains that accumulated in S phase or G2 phase of the cell cycle following gene-product depletion by promoter shut off (Yu et al. 2006). Spontaneous DNA damage was mea- by the relocalization of the DNA damage checkpoint Enrichment analyses S. cerevisiae chromosomes were broken into 5-kb bins. For each bin, the presence or absence of breakpoints and geno- mic features was tabulated. Various genomic features (Di Rienzi et al. 2009) and replication termination sites (Fachinetti et al. 2010) from previous datasets were used for analysis. For each feature, the total number of bins with both the feature and a breakpoint was determined. To test for enrichment of breakpoints and each feature, a hypergeometric distribu- tion was assumed. P-values <0.05 were considered as ev- idence of a correlation and P-values <0.05 after a false sured protein Ddc2 from a diffuse nuclear pattern to discrete sub- nuclear foci (Figure 1A) (Melo et al. 2001; Lisby et al. 2004). Following growth of these strains in doxycycline to repress essential gene expression, the fraction of cells with Ddc2 foci was quantified (Supporting Information, Table S1). We Essential Genome Stability Genes 149 B 50 MAT tester A MAT tester Figure 2 Depletion of yeast es- sential genes results in elevated levels of illegitimate mating. (A) MATa Tet alleles were grown on YPD or YPD containing doxy- cycline (10 ug/ml) for 24 hr and a standard mating test was per- formed using MATa and MATa tester strains. Representative images of strains with elevated levels of illegitimate diploid for- mation following growth in doxycycline are shown. (B) The resulting number of illegitimate diploid colonies that grew with- out doxycycline treatment was subtracted from the number that N 57 N 30 N 35 N 86 NSE1 NUF2 POL2 40 POL30 SPT16 UBC9 wildtype CDC45 MCMS MCM7 MCM4 CSE1 DNA2 DPB11 wildtype >10 1-10 inviable grew with doxycycline treatment and was used to subcategorize For Number of illegitimate diploid colonies +doxycycline (promoter OFF) doxусуcine (promoter ON) the strains into four groups. each group, the distribution of percentage of budded cells with Ddc2-YFP foci was plotted. Bold 15 lines represent the median val- ues, boxes represent the upper and lower quartiles, whiskers represent 1.5 times the inter- quartile range, and outliers are indicated by circles. (C) Compar- ison of Tet alleles with elevated levels of Ddc2 foci and >10 ille- gitimate mating diploid colonies. COC45 NUF2 CSE1 POL2 DNA2 POL30 DPB11 RFC2 MCM4 RFC5 MCM5 SPT16 MCM7 UBC9 42 32 elevated Ddc2 foci 10 illegitimate diploid colonies NSE1 determined that the individual depletion of 47 essential gene products caused an increase of Ddc2 foci relative to wild-type levels, using a cutoff of three standard devia- tions from the wild-type mean (Figure 1B). The gene on tology (GO) processes of the essential genes that were identified are varied (Table S2), but on average the highest levels of Ddc2 foci were observed following the depletion of gene products involved in DNA replication, response to DNA damage stimuli, and cell cycle progression, indicating the im portance of these essential processes in the maintenance of genome integrity (Figure 1B). In addition to the identification of essential genes with defined roles in genome maintenance, 20 essential genes with previously unrecognized contributions to the suppression of spontaneous DNA damage were also identified (Figure 1B, gray bars). Tet allele strains. Mutation or deletion of the MATa locus on chromosome III in haploid cells results in a reversion to the default MATa mating type, termed a-like fakers, allow ing these MATa cells to mate illegitimately with strains of the MATa mating type (Strathern et al. 1981). We determined the levels of a-like faker formation using a patch mating assay (Figure 2A). We found that the depletion of 92 essential genes caused elevated illegitimate mating frequencies both relative to the minus doxycycline control and relative to the wild-type control, indicating loss of genetic information at the MAT locus in these in the presence of doxycycline and 9 strains could not be constructed with the MATa mating type and therefore could not be evaluated. Strains were subcategorized into groups with high (>10 colonies; 57 strains), moderate (1-10 colo- nies; 35 strains), or wild-type (0 colonies; 86 strains) levels of illegitimate mating and the distributions of Ddc2 foci forma- tion for each category were compared (Figure 2B). Both the high and medium categories had greater Ddc2 foci forma- tion when compared to the willd-type category (P-value of 0.022 for high vs. wild type and P-value of 0.028 for me- dium vs. wild type; one-sided Mann-Whitney test), indicat- ing a relationship between the extents of Ddc2 focus formation and the illegitimate mating phenotype. Addi- tionally, strains with spontaneous Ddc2 foci formation above our cutoff were more likely to have increased illegit- imate mating (P-value of 0.00073; hypergeometric test), (Table S3). Thirty strains did not form colonies Depletion of essential gene products causes chromosome loss and rearrangement Increased levels of Ddc2 foci could reflect increased sponta- neous DNA damage, defective repair of spontaneous DNA damage, or a combination of both. An increase in spontaneous DNA damage may not impact genome integrity if the damage is repaired accurately. To directly identify essential genes that suppress chromosome rearrangements and chromosome loss, we used an illegitimate mating assay (Strathern et al. 1981 Lemoine et al. 2005, 2008) that measures loss of genetic information from chromosome III to screen the same 217 E. Cheng et al. 150 oudded cells with Ddc2 foci .
DIC Ddc2-YFP DIC Ddc2-YFP wildtype DPB11 NSE1 Figure 1 Depletion of yeast essential genes results in el- evated levels of spontaneous Ddc2 foci formation. (A) A total of 217 Tet alleles that express Ddc2-YFP and display a G2/M or S phase cell cycle arrest phenotype were grown in the presence of doxycycline (10 ug/ml) for 4 hr to inhibit the transcription of each essential gene. Representative DIC and YFP images are shown for the wild-type, DPB11 and NSE1 strains. Ddc2-YFP foci are indicated with white -doxycycline (promoter ON) +doxycycline (promoter OFF) 3 45 DNA replication Response to DNA damage Cell Cycle Unknown roles in genome maintenance arrows. (B) The percentage of cells with Ddc2-YFP foci is plotted for 47 Tet alleles that showed an increase in Ddc2 foci of at least three standard deviations above the aver- observed in wild type. Bars are shaded according to the GO process annotation of each gene of interest 20 10 5 Tet alleles discovery rate (FDR) correction were considered strongly significant. Restriction digestion and sequencing analysis of FS1 and FS2 Genomic DNA was isolated (Qiagen) from wild-type strains R1158 and BY4741 and digested with EcoRI and Xbal (New England Biolabs) using the suggested conditions. Digested fragments were separated on a 1% agarose gel and hybrid- ized with FEN2 and FS2-2 probes for Southern blot anal- ysis (Table S7). 5' and 3' ends of fragile site 1 (FS1) and FS2 were PCR amplified and sequenced. PCR primers used for both amplification and sequencing are listed in Table S8. Results Depletion of essential gene products causes spontaneous DNA damage We used a collection of tetracycline-regulated promoter alleles (Tet alleles) (Mnaimneh et al. 2004; Yu et al. 2006) of essential genes to systematically identify genes that sup- press spontaneous DNA damage. Since elevated levels of spontaneous DNA damage should elicit a checkpoint re- sponse and cause cell cycle delay, we screened the 217 strains that accumulated in S phase or G2 phase of the cell cycle following gene-product depletion by promoter shut off (Yu et al. 2006). Spontaneous DNA damage was mea- by the relocalization of the DNA damage checkpoint Enrichment analyses S. cerevisiae chromosomes were broken into 5-kb bins. For each bin, the presence or absence of breakpoints and geno- mic features was tabulated. Various genomic features (Di Rienzi et al. 2009) and replication termination sites (Fachinetti et al. 2010) from previous datasets were used for analysis. For each feature, the total number of bins with both the feature and a breakpoint was determined. To test for enrichment of breakpoints and each feature, a hypergeometric distribu- tion was assumed. P-values <0.05 were considered as ev- idence of a correlation and P-values <0.05 after a false sured protein Ddc2 from a diffuse nuclear pattern to discrete sub- nuclear foci (Figure 1A) (Melo et al. 2001; Lisby et al. 2004). Following growth of these strains in doxycycline to repress essential gene expression, the fraction of cells with Ddc2 foci was quantified (Supporting Information, Table S1). We Essential Genome Stability Genes 149 B 50 MAT tester A MAT tester Figure 2 Depletion of yeast es- sential genes results in elevated levels of illegitimate mating. (A) MATa Tet alleles were grown on YPD or YPD containing doxy- cycline (10 ug/ml) for 24 hr and a standard mating test was per- formed using MATa and MATa tester strains. Representative images of strains with elevated levels of illegitimate diploid for- mation following growth in doxycycline are shown. (B) The resulting number of illegitimate diploid colonies that grew with- out doxycycline treatment was subtracted from the number that N 57 N 30 N 35 N 86 NSE1 NUF2 POL2 40 POL30 SPT16 UBC9 wildtype CDC45 MCMS MCM7 MCM4 CSE1 DNA2 DPB11 wildtype >10 1-10 inviable grew with doxycycline treatment and was used to subcategorize For Number of illegitimate diploid colonies +doxycycline (promoter OFF) doxусуcine (promoter ON) the strains into four groups. each group, the distribution of percentage of budded cells with Ddc2-YFP foci was plotted. Bold 15 lines represent the median val- ues, boxes represent the upper and lower quartiles, whiskers represent 1.5 times the inter- quartile range, and outliers are indicated by circles. (C) Compar- ison of Tet alleles with elevated levels of Ddc2 foci and >10 ille- gitimate mating diploid colonies. COC45 NUF2 CSE1 POL2 DNA2 POL30 DPB11 RFC2 MCM4 RFC5 MCM5 SPT16 MCM7 UBC9 42 32 elevated Ddc2 foci 10 illegitimate diploid colonies NSE1 determined that the individual depletion of 47 essential gene products caused an increase of Ddc2 foci relative to wild-type levels, using a cutoff of three standard devia- tions from the wild-type mean (Figure 1B). The gene on tology (GO) processes of the essential genes that were identified are varied (Table S2), but on average the highest levels of Ddc2 foci were observed following the depletion of gene products involved in DNA replication, response to DNA damage stimuli, and cell cycle progression, indicating the im portance of these essential processes in the maintenance of genome integrity (Figure 1B). In addition to the identification of essential genes with defined roles in genome maintenance, 20 essential genes with previously unrecognized contributions to the suppression of spontaneous DNA damage were also identified (Figure 1B, gray bars). Tet allele strains. Mutation or deletion of the MATa locus on chromosome III in haploid cells results in a reversion to the default MATa mating type, termed a-like fakers, allow ing these MATa cells to mate illegitimately with strains of the MATa mating type (Strathern et al. 1981). We determined the levels of a-like faker formation using a patch mating assay (Figure 2A). We found that the depletion of 92 essential genes caused elevated illegitimate mating frequencies both relative to the minus doxycycline control and relative to the wild-type control, indicating loss of genetic information at the MAT locus in these in the presence of doxycycline and 9 strains could not be constructed with the MATa mating type and therefore could not be evaluated. Strains were subcategorized into groups with high (>10 colonies; 57 strains), moderate (1-10 colo- nies; 35 strains), or wild-type (0 colonies; 86 strains) levels of illegitimate mating and the distributions of Ddc2 foci forma- tion for each category were compared (Figure 2B). Both the high and medium categories had greater Ddc2 foci forma- tion when compared to the willd-type category (P-value of 0.022 for high vs. wild type and P-value of 0.028 for me- dium vs. wild type; one-sided Mann-Whitney test), indicat- ing a relationship between the extents of Ddc2 focus formation and the illegitimate mating phenotype. Addi- tionally, strains with spontaneous Ddc2 foci formation above our cutoff were more likely to have increased illegit- imate mating (P-value of 0.00073; hypergeometric test), (Table S3). Thirty strains did not form colonies Depletion of essential gene products causes chromosome loss and rearrangement Increased levels of Ddc2 foci could reflect increased sponta- neous DNA damage, defective repair of spontaneous DNA damage, or a combination of both. An increase in spontaneous DNA damage may not impact genome integrity if the damage is repaired accurately. To directly identify essential genes that suppress chromosome rearrangements and chromosome loss, we used an illegitimate mating assay (Strathern et al. 1981 Lemoine et al. 2005, 2008) that measures loss of genetic information from chromosome III to screen the same 217 E. Cheng et al. 150 oudded cells with Ddc2 foci .

Human Heredity: Principles and Issues (MindTap Course List)
11th Edition
ISBN:9781305251052
Author:Michael Cummings
Publisher:Michael Cummings
Chapter7: Development And Sex Determination
Section: Chapter Questions
Problem 18QP: Equalizing the Expression of X Chromosome Genes in Males and Females Males have only one X...

Related questions

Topic Video
Question
what does the Depletion of essential gene products causes spontaneous DNA damage major result mean in this experiment? (interpretation)
Transcribed Image Text:DIC
Ddc2-YFP
DIC
Ddc2-YFP
wildtype
DPB11
NSE1
Figure 1 Depletion of yeast essential genes results in el-
evated levels of spontaneous Ddc2 foci formation. (A) A
total of 217 Tet alleles that express Ddc2-YFP and display
a G2/M or S phase cell cycle arrest phenotype were grown
in the presence of doxycycline (10 ug/ml) for 4 hr to inhibit
the transcription of each essential gene. Representative
DIC and YFP images are shown for the wild-type, DPB11
and NSE1 strains. Ddc2-YFP foci are indicated with white
-doxycycline
(promoter ON)
+doxycycline
(promoter OFF)
3 45
DNA replication
Response to DNA damage
Cell Cycle
Unknown roles in genome maintenance
arrows. (B) The percentage of cells with Ddc2-YFP foci is
plotted for 47 Tet alleles that showed an increase in Ddc2
foci of at least three standard deviations above the aver-
observed in wild type. Bars are shaded according to
the GO process annotation of each gene of interest
20
10
5
Tet alleles
discovery rate (FDR) correction were considered strongly
significant.
Restriction digestion and sequencing analysis
of FS1 and FS2
Genomic DNA was isolated (Qiagen) from wild-type strains
R1158 and BY4741 and digested with EcoRI and Xbal (New
England Biolabs) using the suggested conditions. Digested
fragments were separated on a 1% agarose gel and hybrid-
ized with FEN2 and FS2-2 probes for Southern blot anal-
ysis (Table S7). 5' and 3' ends of fragile site 1 (FS1) and
FS2 were PCR amplified and sequenced. PCR primers used for
both amplification and sequencing are listed in Table S8.
Results
Depletion of essential gene products causes
spontaneous DNA damage
We used a collection of tetracycline-regulated promoter
alleles (Tet alleles) (Mnaimneh et al. 2004; Yu et al. 2006)
of essential genes to systematically identify genes that sup-
press spontaneous DNA damage. Since elevated levels of
spontaneous DNA damage should elicit a checkpoint re-
sponse and cause cell cycle delay, we screened the 217
strains that accumulated in S phase or G2 phase of the cell
cycle following gene-product depletion by promoter shut
off (Yu et al. 2006). Spontaneous DNA damage was mea-
by the relocalization of the DNA damage checkpoint
Enrichment analyses
S. cerevisiae chromosomes were broken into 5-kb bins. For
each bin, the presence or absence of breakpoints and geno-
mic features was tabulated. Various genomic features (Di
Rienzi et al. 2009) and replication termination sites (Fachinetti
et al. 2010) from previous datasets were used for analysis. For
each feature, the total number of bins with both the feature
and a breakpoint was determined. To test for enrichment of
breakpoints and each feature, a hypergeometric distribu-
tion was assumed. P-values <0.05 were considered as ev-
idence of a correlation and P-values <0.05 after a false
sured
protein Ddc2 from a diffuse nuclear pattern to discrete sub-
nuclear foci (Figure 1A) (Melo et al. 2001; Lisby et al. 2004).
Following growth of these strains in doxycycline to repress
essential gene expression, the fraction of cells with Ddc2 foci
was quantified (Supporting Information, Table S1). We
Essential Genome Stability Genes
149
Transcribed Image Text:B 50
MAT tester
A MAT tester
Figure 2 Depletion of yeast es-
sential genes results in elevated
levels of illegitimate mating. (A)
MATa Tet alleles were grown
on YPD or YPD containing doxy-
cycline (10 ug/ml) for 24 hr and
a standard mating test was per-
formed using MATa and MATa
tester strains. Representative
images of strains with elevated
levels of illegitimate diploid for-
mation following growth in
doxycycline are shown. (B) The
resulting number of illegitimate
diploid colonies that grew with-
out doxycycline treatment was
subtracted from the number that
N 57
N 30
N 35
N 86
NSE1
NUF2
POL2
40
POL30
SPT16
UBC9
wildtype
CDC45
MCMS
MCM7
MCM4
CSE1
DNA2
DPB11
wildtype
>10
1-10
inviable
grew with doxycycline treatment
and was used to subcategorize
For
Number of illegitimate diploid colonies
+doxycycline
(promoter OFF)
doxусуcine
(promoter ON)
the strains into four groups.
each group, the distribution of
percentage of budded cells with
Ddc2-YFP foci was plotted. Bold
15
lines represent the median val-
ues, boxes represent the upper
and lower quartiles, whiskers
represent 1.5 times the inter-
quartile range, and outliers are
indicated by circles. (C) Compar-
ison of Tet alleles with elevated
levels of Ddc2 foci and >10 ille-
gitimate mating diploid colonies.
COC45 NUF2
CSE1 POL2
DNA2 POL30
DPB11 RFC2
MCM4 RFC5
MCM5 SPT16
MCM7 UBC9
42
32
elevated Ddc2
foci
10 illegitimate
diploid colonies
NSE1
determined that the individual depletion of 47 essential
gene products caused an increase of Ddc2 foci relative to
wild-type levels, using a cutoff of three standard devia-
tions from the wild-type mean (Figure 1B). The gene on
tology (GO) processes of the essential genes that were
identified are varied (Table S2), but on average the highest
levels of Ddc2 foci were observed following the depletion of
gene products involved in DNA replication, response to DNA
damage stimuli, and cell cycle progression, indicating the im
portance of these essential processes in the maintenance of
genome integrity (Figure 1B). In addition to the identification
of essential genes with defined roles in genome maintenance,
20 essential genes with previously unrecognized contributions
to the suppression of spontaneous DNA damage were also
identified (Figure 1B, gray bars).
Tet allele strains. Mutation or deletion of the MATa locus
on chromosome III in haploid cells results in a reversion to
the default MATa mating type, termed a-like fakers, allow
ing these MATa cells to mate illegitimately with strains of
the MATa mating type (Strathern et al. 1981). We determined
the levels of a-like faker formation using a patch mating assay
(Figure 2A). We found that the depletion of 92 essential genes
caused elevated illegitimate mating frequencies both relative
to the minus doxycycline control and relative to the wild-type
control, indicating loss of genetic information at the MAT locus
in these
in the presence of doxycycline and 9 strains could not be
constructed with the MATa mating type and therefore could
not be evaluated. Strains were subcategorized into groups
with high (>10 colonies; 57 strains), moderate (1-10 colo-
nies; 35 strains), or wild-type (0 colonies; 86 strains) levels of
illegitimate mating and the distributions of Ddc2 foci forma-
tion for each category were compared (Figure 2B). Both the
high and medium categories had greater Ddc2 foci forma-
tion when compared to the willd-type category (P-value of
0.022 for high vs. wild type and P-value of 0.028 for me-
dium vs. wild type; one-sided Mann-Whitney test), indicat-
ing a relationship between the extents of Ddc2 focus
formation and the illegitimate mating phenotype. Addi-
tionally, strains with spontaneous Ddc2 foci formation
above our cutoff were more likely to have increased illegit-
imate mating (P-value of 0.00073; hypergeometric test),
(Table S3). Thirty strains did not form colonies
Depletion of essential gene products causes
chromosome loss and rearrangement
Increased levels of Ddc2 foci could reflect increased sponta-
neous DNA damage, defective repair of spontaneous DNA
damage, or a combination of both. An increase in spontaneous
DNA damage may not impact genome integrity if the damage
is repaired accurately. To directly identify essential genes that
suppress chromosome rearrangements and chromosome loss,
we used an illegitimate mating assay (Strathern et al. 1981
Lemoine et al. 2005, 2008) that measures loss of genetic
information from chromosome III to screen the same 217
E. Cheng et al.
150
oudded cells with Ddc2 foci
.
Expert Solution
This question has been solved!
Explore an expertly crafted, step-by-step solution for a thorough understanding of key concepts.

Step by step
Solved in 3 steps

Knowledge Booster


Learn more about
Need a deep-dive on the concept behind this application? Look no further. Learn more about this topic, biology and related others by exploring similar questions and additional content below.Recommended textbooks for you
Human Heredity: Principles and Issues (MindTap Co…
Biology
ISBN:
9781305251052
Author:
Michael Cummings
Publisher:
Cengage Learning
Human Heredity: Principles and Issues (MindTap Co…
Biology
ISBN:
9781305251052
Author:
Michael Cummings
Publisher:
Cengage Learning