Please summarize these papers and explain what the figures mean. Please elaborate as if your explaining the class what this paper is about. Please explain what the figure tells us and what they do.
Please summarize these papers and explain what the figures mean. Please elaborate as if your explaining the class what this paper is about. Please explain what the figure tells us and what they do.

Biology 2e
2nd Edition
ISBN:9781947172517
Author:Matthew Douglas, Jung Choi, Mary Ann Clark
Publisher:Matthew Douglas, Jung Choi, Mary Ann Clark
Chapter9: Cell Communication
Section: Chapter Questions
Problem 12RQ: Histamine binds to the H1 G-protein-linked receptor to initiate the itchiness and airway...

Related questions
Question
Please summarize these papers and explain what the figures mean. Please elaborate as if your explaining the class what this paper is about. Please explain what the figure tells us and what they do.
![3.5
3.0
2.5
2.0
1.5
1.0
e 3.0
2.5
[Ca], (a.u.)
2.0
0
1.5
1.0
ли
6 8
Time (min)
Ouabain
2
0
4
MULT
Na,K-ATPase and InsP3 Receptor in a Signaling Microdomain
b
20
10 12
2-APB
60
f
80
3.0
2.5
2.0
1.5
1.0
[Ca], (a.u.)
3.5
3.0
2.5
2.0
1.5
1.0
um
10
Time (min)
0
0
5
5
15
20
nounced increase of basal cytosolic Ca2+, consistent with
inhibition of the SERCA pump (19). Since available data sug-
gest that low concentrations (up to 20 μM) of 2-APB will pref-
erentially inhibit InsP³Rs, we tested the effect of 5 μM 2-APB.
Using this concentration, we found that ouabain-induced Ca²+
oscillations were abolished in the majority of cells treated with
2-APB (5 μM) (Fig. 1e). Collectively, the inhibitory effects of
CPA and 2-APB demonstrate that release of Ca²+ via InsP3R is
an essential contributor to the Ca²+ oscillations triggered by
the ouabain/Na,K-ATPase complex.
C 2.5
Activation of InsP3Rs is critically dependent on activation of
phospholipase C (PLC), phosphatidylinositol lipid hydrolysis,
and liberation of InsP3. Notably, however, recent studies indi-
cate that InsP3R function is also modulated by interaction with
accessory proteins (20-22). To examine the role of InsP3 for the
ouabain-induced Ca²+ oscillations, RPT cells were transfected
with a construct encoding a hyper-affinity InsP3 absorbent, an
InsP3 sponge. The InsP3 sponge, having more than 1000-fold
higher affinity for InsP; than InsP³R, traps InsP3 and abro-
gates InsP3-induced Ca²+ release (10). The construct also en-
coded GFP to facilitate identification of transfected cells. Oua-
bain triggered low frequency Ca2+ oscillations in one-third of
the cells expressing the InsP3 sponge (Fig. 1f). The amplitude
of the oscillatory response was attenuated in some, but not all,
of the cells expressing the InsP3 sponge. To confirm the effi-
ciency of the InsP3 sponge in quenching InsP3-mediated Ca²+
signaling in RPT cells, we treated cells with bradykinin, a well
known activator of PLC and InsP3 production (23). Bradykinin
induced single Ca²+ transients in virtually all non-transfected
cells but was without effect in all cells expressing the InsP3
sponge (Fig. 1g). Cells expressing only GFP exhibited regular
ouabain-induced Ca²+ oscillations (data not shown). It was
further found that preincubation of RPT cells with a PLC
inhibitor, U73122, abolished bradykinin-induced Ca²+ tran-
sients (data not shown) but did not influence ouabain-induced
Ca²+ oscillations (Fig. 1h). These findings indicate that
ouabain-induced Ca²+ oscillations do not require increased
InsP3 levels to activate InsP3R in this model.
Immunocytochemical studies, performed on COS-7 cells, re-
vealed partial co-localization of Na,K-ATPase with InsP3R
types 1, 2, and 3 (InsP3R1, InsP3R2, and InsP3R3), respec-
tively. Only InsP3R2 (Fig. 2a) and InsP3R3 (Fig. 2b) were
studied in subsequent experiments since these isoforms were
more abundantly expressed than InsP³R1. To investigate the
E
2.0
1.5
[Ca], (a.u.)
1.0
9 3.5
3.0
2.5
2.0
1.5
1.0
0
-5
20 40
Time (min)
М
Add
40
10 15 20 25 30
Time (min)
5
Time (min)
Time (min)
FIG. 1. Intracellular Ca²+ response to ouabain in renal cells. a-c, Ca²+ oscillations in 250 μM ouabain-treated RPT cells (a), 250 nm
ouabain-treated COS-7 cells (b), and 100 pM ouabain-treated RPT cells (c). Arbitrary units (a.u.) represent ratio values corresponding to
intracellular Ca²+ concentration changes. In d, CPA depleted intracellular ER Ca²+ stores and abolished ouabain-induced Ca²+ oscillations in RPT
cells. Each trace represents a single cell recording. In e, ouabain-induced Ca²+ oscillations in RPT cells were abolished by 2-APB (5 μm). f, two
representative single cell recordings of cytosolic Ca²+ in RPT cells transfected with the InsP, sponge (n = 15). The amplitude was lower in some
InsP3 sponge-expressing cells. g, two representative single cell recordings of cytosolic Ca²+ in RPT cells. Bradykinin did not induce Ca²+ transients
in InsP, sponge-expressing RPT cells (trace A), whereas non-transfected cells exhibited Ca²+ transients (trace B). In h, U73122 (5 µM) did not
abolish ouabain-induced Ca²+ oscillations in RPT cells.
0
GFP-NKAα1
60 80
10
d
[Ca], (a.u.) P
15
h
4.0
[Ca], (a.u.)
3.0
2.0
1.0
3.5
3.0
2.5
2.0
1.5
1.0
0
InsP₂R2-Cy3
10
0
CPA
3
Ouabain
20 30 40 50
Time (min)
U73122
Ouabain
50357
л
6
9
Time (min)
Merged
12 15
GFP-NKA 1
InsP-R3-Cy3
Merged
FIG. 2. Immunocytochemical studies of Na,K-ATPase and
InsP,R localization in COS-7 cells. Na,K-ATPase (GFP-NKAa1) and
InsP,R2 (InsP.R2-Cy3) (a) or InsP.R3 (InsP.R3-Cy3) (6) co-localize
near the plasma membrane.
spatial relationship between Na,K-ATPase and InsP³R on a
nanometer scale, FRET measurements were performed. In this
protocol, we used COS-7 cells stably expressing GFP-tagged
Na,K-ATPase al-subunit. These cells express approximately
the same level of Na,K-ATPase as wild type COS-7 cells (12).
GFP, which was fused to the cytosolic NH₂ terminus of Na,K-
ATPase, served as FRET donor (GFP-NKAa1). The primary
antibodies against InsP3R2 or InsP3R3 were probed with a
Cy3-conjugated IgG secondary antibody, which served as the
FRET acceptor (InsP3R-Cy3). The epitopes recognized by the
InsP3R2 and InsP3R3 antibodies are located in the cytoplasmic
COOH terminus of the respective InsP3Rs (13). The GFP-
NKAa1 fluorescence intensity was, following acceptor photo-
bleaching, enhanced 12.5 ± 0.9% for InsP3R2 and 15.5 ± 2.0%
for InsP3R3 (Fig. 3, a and b). These results imply that the donor
and acceptor complexes, GFP-NKAa1 and anti-InsP3R-anti-
mouse IgG-Cy3, were separated less than 12 nm, i.e. the max-
imal distance for FRET detection between GFP and Cy3 (16).
Ouabain treatment significantly increased FRET (from 15.5 ±
2.0% to 25.0± 1.6%) between Na,K-ATPase and InsP,R (Fig. 3,
a and b).
To confirm that the observed FRET between Na,K-ATPase
and InsP3R3 was a unique property of this pair of proteins and
not merely the result of non-specific experimental artifacts, we](/v2/_next/image?url=https%3A%2F%2Fcontent.bartleby.com%2Fqna-images%2Fquestion%2F10b2558f-14bb-4a79-9eba-8554e0210a1c%2Ff42f366a-aa83-45d5-b148-d0adeb125923%2Face3b8m_processed.jpeg&w=3840&q=75)
Transcribed Image Text:3.5
3.0
2.5
2.0
1.5
1.0
e 3.0
2.5
[Ca], (a.u.)
2.0
0
1.5
1.0
ли
6 8
Time (min)
Ouabain
2
0
4
MULT
Na,K-ATPase and InsP3 Receptor in a Signaling Microdomain
b
20
10 12
2-APB
60
f
80
3.0
2.5
2.0
1.5
1.0
[Ca], (a.u.)
3.5
3.0
2.5
2.0
1.5
1.0
um
10
Time (min)
0
0
5
5
15
20
nounced increase of basal cytosolic Ca2+, consistent with
inhibition of the SERCA pump (19). Since available data sug-
gest that low concentrations (up to 20 μM) of 2-APB will pref-
erentially inhibit InsP³Rs, we tested the effect of 5 μM 2-APB.
Using this concentration, we found that ouabain-induced Ca²+
oscillations were abolished in the majority of cells treated with
2-APB (5 μM) (Fig. 1e). Collectively, the inhibitory effects of
CPA and 2-APB demonstrate that release of Ca²+ via InsP3R is
an essential contributor to the Ca²+ oscillations triggered by
the ouabain/Na,K-ATPase complex.
C 2.5
Activation of InsP3Rs is critically dependent on activation of
phospholipase C (PLC), phosphatidylinositol lipid hydrolysis,
and liberation of InsP3. Notably, however, recent studies indi-
cate that InsP3R function is also modulated by interaction with
accessory proteins (20-22). To examine the role of InsP3 for the
ouabain-induced Ca²+ oscillations, RPT cells were transfected
with a construct encoding a hyper-affinity InsP3 absorbent, an
InsP3 sponge. The InsP3 sponge, having more than 1000-fold
higher affinity for InsP; than InsP³R, traps InsP3 and abro-
gates InsP3-induced Ca²+ release (10). The construct also en-
coded GFP to facilitate identification of transfected cells. Oua-
bain triggered low frequency Ca2+ oscillations in one-third of
the cells expressing the InsP3 sponge (Fig. 1f). The amplitude
of the oscillatory response was attenuated in some, but not all,
of the cells expressing the InsP3 sponge. To confirm the effi-
ciency of the InsP3 sponge in quenching InsP3-mediated Ca²+
signaling in RPT cells, we treated cells with bradykinin, a well
known activator of PLC and InsP3 production (23). Bradykinin
induced single Ca²+ transients in virtually all non-transfected
cells but was without effect in all cells expressing the InsP3
sponge (Fig. 1g). Cells expressing only GFP exhibited regular
ouabain-induced Ca²+ oscillations (data not shown). It was
further found that preincubation of RPT cells with a PLC
inhibitor, U73122, abolished bradykinin-induced Ca²+ tran-
sients (data not shown) but did not influence ouabain-induced
Ca²+ oscillations (Fig. 1h). These findings indicate that
ouabain-induced Ca²+ oscillations do not require increased
InsP3 levels to activate InsP3R in this model.
Immunocytochemical studies, performed on COS-7 cells, re-
vealed partial co-localization of Na,K-ATPase with InsP3R
types 1, 2, and 3 (InsP3R1, InsP3R2, and InsP3R3), respec-
tively. Only InsP3R2 (Fig. 2a) and InsP3R3 (Fig. 2b) were
studied in subsequent experiments since these isoforms were
more abundantly expressed than InsP³R1. To investigate the
E
2.0
1.5
[Ca], (a.u.)
1.0
9 3.5
3.0
2.5
2.0
1.5
1.0
0
-5
20 40
Time (min)
М
Add
40
10 15 20 25 30
Time (min)
5
Time (min)
Time (min)
FIG. 1. Intracellular Ca²+ response to ouabain in renal cells. a-c, Ca²+ oscillations in 250 μM ouabain-treated RPT cells (a), 250 nm
ouabain-treated COS-7 cells (b), and 100 pM ouabain-treated RPT cells (c). Arbitrary units (a.u.) represent ratio values corresponding to
intracellular Ca²+ concentration changes. In d, CPA depleted intracellular ER Ca²+ stores and abolished ouabain-induced Ca²+ oscillations in RPT
cells. Each trace represents a single cell recording. In e, ouabain-induced Ca²+ oscillations in RPT cells were abolished by 2-APB (5 μm). f, two
representative single cell recordings of cytosolic Ca²+ in RPT cells transfected with the InsP, sponge (n = 15). The amplitude was lower in some
InsP3 sponge-expressing cells. g, two representative single cell recordings of cytosolic Ca²+ in RPT cells. Bradykinin did not induce Ca²+ transients
in InsP, sponge-expressing RPT cells (trace A), whereas non-transfected cells exhibited Ca²+ transients (trace B). In h, U73122 (5 µM) did not
abolish ouabain-induced Ca²+ oscillations in RPT cells.
0
GFP-NKAα1
60 80
10
d
[Ca], (a.u.) P
15
h
4.0
[Ca], (a.u.)
3.0
2.0
1.0
3.5
3.0
2.5
2.0
1.5
1.0
0
InsP₂R2-Cy3
10
0
CPA
3
Ouabain
20 30 40 50
Time (min)
U73122
Ouabain
50357
л
6
9
Time (min)
Merged
12 15
GFP-NKA 1
InsP-R3-Cy3
Merged
FIG. 2. Immunocytochemical studies of Na,K-ATPase and
InsP,R localization in COS-7 cells. Na,K-ATPase (GFP-NKAa1) and
InsP,R2 (InsP.R2-Cy3) (a) or InsP.R3 (InsP.R3-Cy3) (6) co-localize
near the plasma membrane.
spatial relationship between Na,K-ATPase and InsP³R on a
nanometer scale, FRET measurements were performed. In this
protocol, we used COS-7 cells stably expressing GFP-tagged
Na,K-ATPase al-subunit. These cells express approximately
the same level of Na,K-ATPase as wild type COS-7 cells (12).
GFP, which was fused to the cytosolic NH₂ terminus of Na,K-
ATPase, served as FRET donor (GFP-NKAa1). The primary
antibodies against InsP3R2 or InsP3R3 were probed with a
Cy3-conjugated IgG secondary antibody, which served as the
FRET acceptor (InsP3R-Cy3). The epitopes recognized by the
InsP3R2 and InsP3R3 antibodies are located in the cytoplasmic
COOH terminus of the respective InsP3Rs (13). The GFP-
NKAa1 fluorescence intensity was, following acceptor photo-
bleaching, enhanced 12.5 ± 0.9% for InsP3R2 and 15.5 ± 2.0%
for InsP3R3 (Fig. 3, a and b). These results imply that the donor
and acceptor complexes, GFP-NKAa1 and anti-InsP3R-anti-
mouse IgG-Cy3, were separated less than 12 nm, i.e. the max-
imal distance for FRET detection between GFP and Cy3 (16).
Ouabain treatment significantly increased FRET (from 15.5 ±
2.0% to 25.0± 1.6%) between Na,K-ATPase and InsP,R (Fig. 3,
a and b).
To confirm that the observed FRET between Na,K-ATPase
and InsP3R3 was a unique property of this pair of proteins and
not merely the result of non-specific experimental artifacts, we
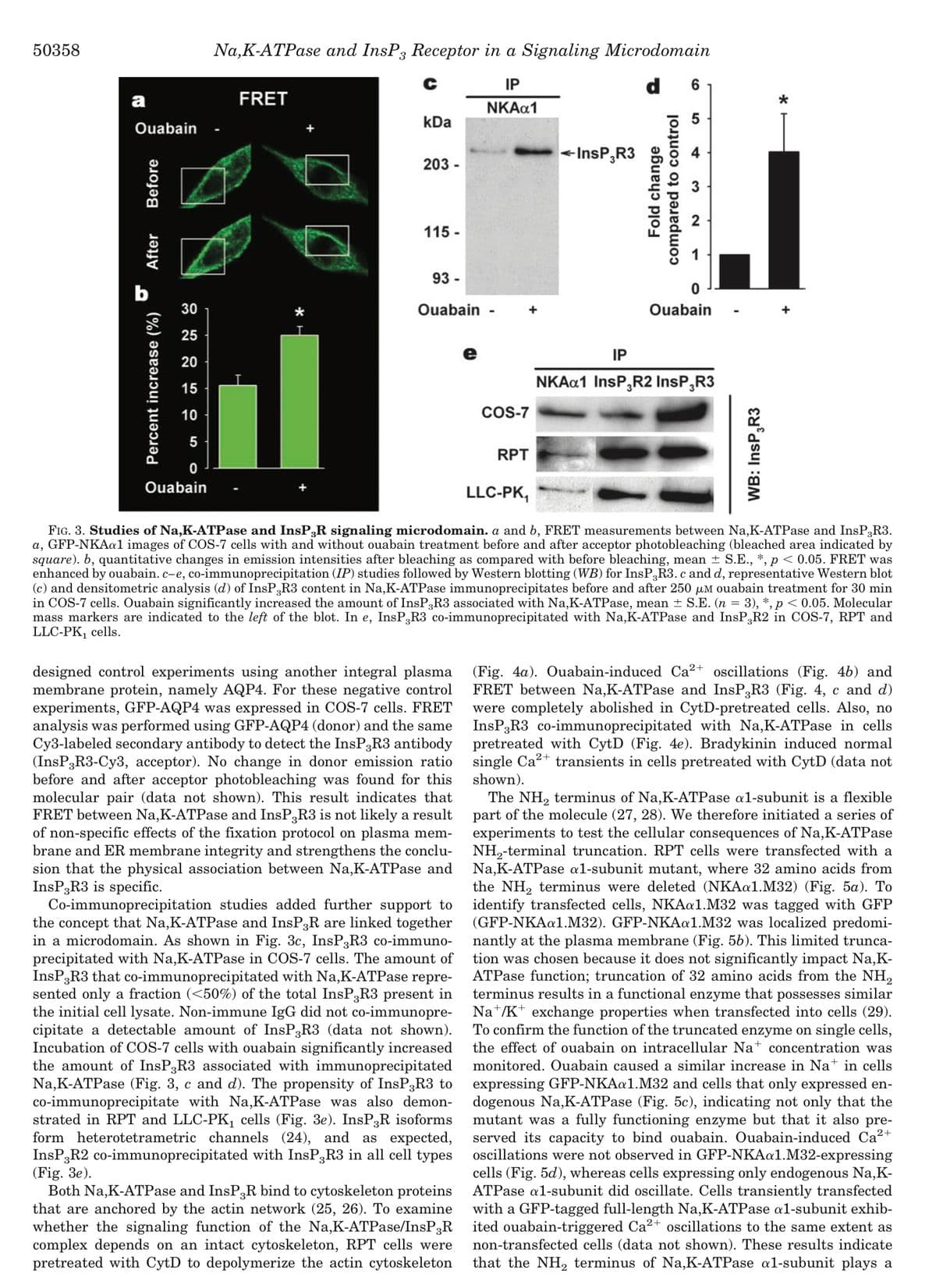
Transcribed Image Text:50358
a
Ouabain
Before
After
30
25
20
15
10
5
0
Ouabain
Percent increase (%)
Na,K-ATPase and InsP3 Receptor in a Signaling Microdomain
IP
d 6
NKAa1
FRET
kDa
203-
115-
93-
Ouabain
designed control experiments using another integral plasma
membrane protein, namely AQP4. For these negative control
experiments, GFP-AQP4 was expressed in COS-7 cells. FRET
analysis was performed using GFP-AQP4 (donor) and the same
Cy3-labeled secondary antibody to detect the InsP3R3 antibody
(InsP3R3-Cy3, acceptor). No change in donor emission ratio
before and after acceptor photobleaching was found for this
molecular pair (data not shown). This result indicates that
FRET between Na,K-ATPase and InsP3R3 is not likely a result
of non-specific effects of the fixation protocol on plasma mem-
brane and ER membrane integrity and strengthens the conclu-
sion that the physical association between Na,K-ATPase and
InsP3R3 is specific.
Co-immunoprecipitation studies added further support to
the concept that Na,K-ATPase and InsP3R are linked together
in a microdomain. As shown in Fig. 3c, InsP3R3 co-immuno-
precipitated with Na,K-ATPase in COS-7 cells. The amount of
InsP3R3 that co-immunoprecipitated with Na,K-ATPase repre-
sented only a fraction (<50%) of the total InsP3R3 present in
the initial cell lysate. Non-immune IgG did not co-immunopre-
cipitate a detectable amount of InsP3R3 (data not shown).
Incubation of COS-7 cells with ouabain significantly increased
the amount of InsP3R3 associated with immunoprecipitated
Na,K-ATPase (Fig. 3, c and d). The propensity of InsP3R3 to
co-immunoprecipitate with Na,K-ATPase was also demon-
strated in RPT and LLC-PK₁ cells (Fig. 3e). InsP3R isoforms
form heterotetrametric channels (24), and as expected,
InsP3R2 co-immunoprecipitated with InsP3R3 in all cell types
(Fig. 3e).
e
Both Na,K-ATPase and InsP3R bind to cytoskeleton proteins
that are anchored by the actin network (25, 26). To examine
whether the signaling function of the Na,K-ATPase/InsP3R
complex depends on an intact cytoskeleton, RPT cells were
pretreated with CytD to depolymerize the actin cytoskeleton
COS-7
RPT
InsP3R3
Fold change
compared to control
LO
5
4
3
N
0
Ouabain
IP
NKAa1 InsP3R2 InsP-R3
LLC-PK₁
FIG. 3. Studies of Na,K-ATPase and InsP;R signaling microdomain. a and b, FRET measurements between Na,K-ATPase and InsP3R3.
a, GFP-NKAa1 images of COS-7 cells with and without ouabain treatment before and after acceptor photobleaching (bleached area indicated by
square). b, quantitative changes in emission intensities after bleaching as compared with before bleaching, mean ± S.E., *, p < 0.05. FRET was
enhanced by ouabain. c-e, co-immunoprecipitation (IP) studies followed by Western blotting (WB) for InsP3R3. c and d, representative Western blot
(c) and densitometric analysis (d) of InsP3R3 content in Na,K-ATPase immunoprecipitates before and after 250 μM ouabain treatment for 30 min
in COS-7 cells. Ouabain significantly increased the amount of InsP3R3 associated with Na,K-ATPase, mean ± S.E. (n = 3), *, p < 0.05. Molecular
mass markers are indicated to the left of the blot. In e, InsP,R3 co-immunoprecipitated with Na,K-ATPase and InsP,R2 in COS-7, RPT and
LLC-PK₁ cells.
WB: InsP3R3
*
(Fig. 4a). Ouabain-induced Ca²+ oscillations (Fig. 4b) and
FRET between Na,K-ATPase and InsP3R3 (Fig. 4, c and d)
were completely abolished in CytD-pretreated cells. Also, no
InsP3R3 co-immunoprecipitated with Na,K-ATPase in cells
pretreated with CytD (Fig. 4e). Bradykinin induced normal
single Ca²+ transients in cells pretreated with CytD (data not
shown).
The NH₂ terminus of Na,K-ATPase a1-subunit is a flexible
part of the molecule (27, 28). We therefore initiated a series of
experiments to test the cellular consequences of Na,K-ATPase
NH₂-terminal truncation. RPT cells were transfected with a
Na,K-ATPase al-subunit mutant, where 32 amino acids from
the NH₂ terminus were deleted (NKAa1.M32) (Fig. 5a). To
identify transfected cells, NKAa1.M32 was tagged with GFP
(GFP-NKAa1.M32). GFP-NKAa1.M32 was localized predomi-
nantly at the plasma membrane (Fig. 56). This limited trunca-
tion was chosen because it does not significantly impact Na,K-
ATPase function; truncation of 32 amino acids from the NH₂
terminus results in a functional enzyme that possesses similar
Na/K+ exchange properties when transfected into cells (29).
To confirm the function of the truncated enzyme on single cells,
the effect of ouabain on intracellular Na+ concentration was
monitored. Ouabain caused a similar increase in Nat in cells
expressing GFP-NKAa1.M32 and cells that only expressed en-
dogenous Na,K-ATPase (Fig. 5c), indicating not only that the
mutant was a fully functioning enzyme but that it also pre-
served its capacity to bind ouabain. Ouabain-induced Ca²+
oscillations were not observed in GFP-NKAa1.M32-expressing
cells (Fig. 5d), whereas cells expressing only endogenous Na,K-
ATPase al-subunit did oscillate. Cells transiently transfected
with a GFP-tagged full-length Na,K-ATPase a1-subunit exhib-
ited ouabain-triggered Ca²+ oscillations to the same extent as
non-transfected cells (data not shown). These results indicate
that the NH₂ terminus of Na,K-ATPase al-subunit plays a
Expert Solution
This question has been solved!
Explore an expertly crafted, step-by-step solution for a thorough understanding of key concepts.

Step by step
Solved in 4 steps

Knowledge Booster


Learn more about
Need a deep-dive on the concept behind this application? Look no further. Learn more about this topic, biology and related others by exploring similar questions and additional content below.Recommended textbooks for you
Biology 2e
Biology
ISBN:
9781947172517
Author:
Matthew Douglas, Jung Choi, Mary Ann Clark
Publisher:
OpenStax
Biology 2e
Biology
ISBN:
9781947172517
Author:
Matthew Douglas, Jung Choi, Mary Ann Clark
Publisher:
OpenStax